付巧妹的博士导师是2022年诺贝尔奖得主斯万特·帕博。
该文发表时间:2023年6月。中文媒体几乎没有报道。
东亚人进入亚洲的事实已无法掩盖。
人类学学报, 2023, 42(03): 412-421 doi: 10.16359/j.1000-3193/AAS.2023.0010
综述
古基因组揭示史前欧亚大陆现代人复杂遗传历史
张明 ,1,2,3, 平婉菁2,3, 付巧妹,2,3
,1,2,3, 平婉菁2,3, 付巧妹,2,3
1.中国-中亚人类与环境“一带一路”联合实验室, 文化遗产研究与保护技术教育部重点实验室, 西北大学文化遗产学院, 西安710127
2.中国科学院古脊椎动物与古人类研究所, 中国科学院脊椎动物演化与人类起源重点实验室, 北京100044
3.中国科学院生物演化与环境卓越创新中心, 北京100044
4.里士满大学生物系, 里士满VA 23173
Ancient genomes reveal the complex genetic history of Prehistoric Eurasian modern humans
ZHANG Ming,1,2,3, PING Wanjing2,3, YANG Melinda Anna2,4, FU Qiaomei,2,3
1. China-Central Asia “the Belt and Road” Joint Laboratory on Human and Environment Research, Key Laboratory of Cultural Heritage Research and Conservation, School of Culture Heritage, Northwest University, Xi’an 710127
2. Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044
3. CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044
4. Department of Biology, University of Richmond, Richmond, VA 23173, USA

古DNA提取与测序技术的发展,让科学家们能够利用史前现代人样本直接开展古基因组研究。古基因组研究发现,在约4万年前,欧亚大陆上至少存在着6个独立的现代人群体,其中3个群体并未对现今人群贡献基因。在距今4万年至末次盛冰期结束,欧亚大陆至少有5个具有代表性的现代人群体。末次盛冰期结束后的部分现代人群体与现今人群拥有更近的遗传关系,而部分群体则未对现今人群贡献基因。本文基于末次盛冰期前(45~19 kaBP)与末次盛冰期后(19~10 kaBP)两个重要时间段的欧亚大陆史前现代人的基因组研究,梳理欧亚现代人在时间与空间上的发展脉络,重点探究此前研究相对滞后的欧亚大陆东部地区。
关键词: 欧亚大陆; 现代人; 古DNA; 人群结构; 遗传历史
Significant shifts in human populations occurred several times throughout history, as populations dispersed throughout Eurasia about 50 kaBP. During the Last Glacial Maximum (LGM), global temperatures dropped sharply causing environmental deterioration and population turnover in areas. After the LGM, populations increased as the natural environment stabilized and gradually developed into today’s populations. With advancements in ancient DNA extraction and sequencing technology, it is increasingly possible to directly retrieve genome-wide data from prehistoric modern human remains. The rapid emergence of new ancient genomes provides an entirely new direction for studying modern human population structure and evolutionary history. This research on Eurasian populations spanning 45~19 kaBP (pre-LGM) and 19~10 kaBP (post-LGM) summarizes the movement and interaction of prehistoric modern human populations, focusing especially on prehistoric East Eurasia, a region that has been less well-studied genetically. Of at least six distinct populations in Eurasia, three did not contribute substantial ancestry to present-day populations: Ust’-Ishim (≈45 kaBP) from northwestern Siberia; Oase 1 (≈40 kaBP) from Romania; and Zlatý k?ň (over 45 kaBP) from Czechia. One population represented by three individuals (4.6~4.3 kaBP, from Bacho Kiro Cave, Bulgaria) seemed to contribute at least a partial genetic component to later some Eurasian populations. One population represented by Tianyuan man (≈40 kaBP, from East Asia) was shown to be more similar to present-day East Asians and Native Americans than to present-day or ancient Europeans. One population represented by Kostenki 14 (≈36 kaBP, from western Siberia) and Goyet Q116-1 (≈35 kaBP, from Belgium) was more closely related to Europeans than to other Eurasians. This work also summarized five representative populations after 40 kaBP and before the end of the LGM. In East Eurasia after the LGM (or since 14 kaBP), population histories played out very differently. For instance, high genetic continuity is observed in the Amur region in the last 14 kaBP, while in the Guangxi region of southern China, an ancient population that lived 10.5 kaBP carried ancestry not represented in any present-day humans. To conclude, comparison of genome-wide ancient DNA from multiple prehistoric humans have illustrated a complex genetic history of prehistoric Eurasian modern humans. In the future, additional ancient genomes will provide more evidence and details to illuminate the complex genetic history of modern humans.
Keywords: Eurasia; modern human; Ancient DNA; Population structure; Genetic history
本文引用格式
张明, 平婉菁, 付巧妹. 古基因组揭示史前欧亚大陆现代人复杂遗传历史[J]. 人类学学报, 2023, 42(03): 412-421 doi:10.16359/j.1000-3193/AAS.2023.0010
ZHANG Ming, PING Wanjing, YANG Melinda Anna, FU Qiaomei. Ancient genomes reveal the complex genetic history of Prehistoric Eurasian modern humans[J]. Acta Anthropologica Sinica, 2023, 42(03): 412-421 doi:10.16359/j.1000-3193/AAS.2023.0010
1 引言
古基因组研究表明,欧亚大陆在史前时期存在多个现代人群体,且这些人群的结构在50 kaBP以来发生了几次重要改变。现代人快速且成功地扩散至整个欧亚大陆乃至于美洲[1⇓-3],可能与这一时期地球气候的几次冷暖更替事件相关[4]。末次盛冰期(约26.5 kaBP开始,一直持续到约19 kaBP)时,全球气温骤降,自然环境极度恶化,一些地区的现代人面临着严峻的生存挑战,有些地区的人群结构可能因此而发生颠覆性的改变,造成冰期后再迁入的人群与之前的人群不存在直接的基因联系[5]。末次盛冰期之后,地球环境进入相对稳定并且适宜人类生存的时期,各地的现代人群体不断发展、迁徙、融合,逐渐形成了现今现代人的群体结构。
近年来,古DNA实验技术与研究方法的高速发展[6⇓⇓-9],极大地推动了人类学的研究。通过古DNA技术直接获取史前材料中的遗传信息,结合材料发现的地点和测年信息,研究者们得以在一个更大的时空框架下开展综合分析。相比于传统的化石形态学研究,古DNA研究对于材料完整度的要求相对较低,并且能够提供更为客观的数据信息。此外,由于每一个个体的古基因组数据都能够在很大程度上反映群体的遗传特征,所以单个个体的古基因组数据能够代表整个群体进行分析,这对于只发现零星人骨遗存的遗址来说,具有更加重要的意义。自2010年,4 kaBP的爱斯基摩人、尼安德特人及丹尼索瓦人的基因组数据发布以来[6,10,11],已经有超过6000例人类古基因组数据发布[12],相关古基因组研究发现和结论已经证实、推翻或修正了古人类学领域许多已有的观点和假说,并提出了许多新的科学认知。如,证实除尼安德特人外还存在另外一种灭绝古人类──丹尼索瓦人[11],发现灭绝古人类(尼安德特人和丹尼索瓦人)与现代人有过基因交流[6,11],推翻了此前认为的现代人与尼安德特人自分离后不再有基因交流的观点;还修正了此前欧亚大陆东部地区的“两层假说”,发现新石器时代早期中国南方福建等沿海地区古人群,实质上并无明显属于“第一层”人群的遗传信息[13]。2017年以前,已发表的古基因组数据主要集中于欧亚大陆西部,科学家们对欧洲、北亚等区域的人群演化问题已展开比较详尽的研究。2017年以来,随着科学家们把关注的重心移向欧亚大陆东部地区,欧亚大陆东部人类古基因组数据开始持续发布,并涌现出了诸多重要的成果,使这一区域人类古基因组研究相对落后的局面已开始逐渐改变。本文重点关注欧亚大陆东部地区史前现代人相关的重要古基因组研究成果,希望为读者梳理、总结欧亚现代人,尤其是欧亚东部现代人在时间与空间上的发展脉络。
2 遗传历史
图1
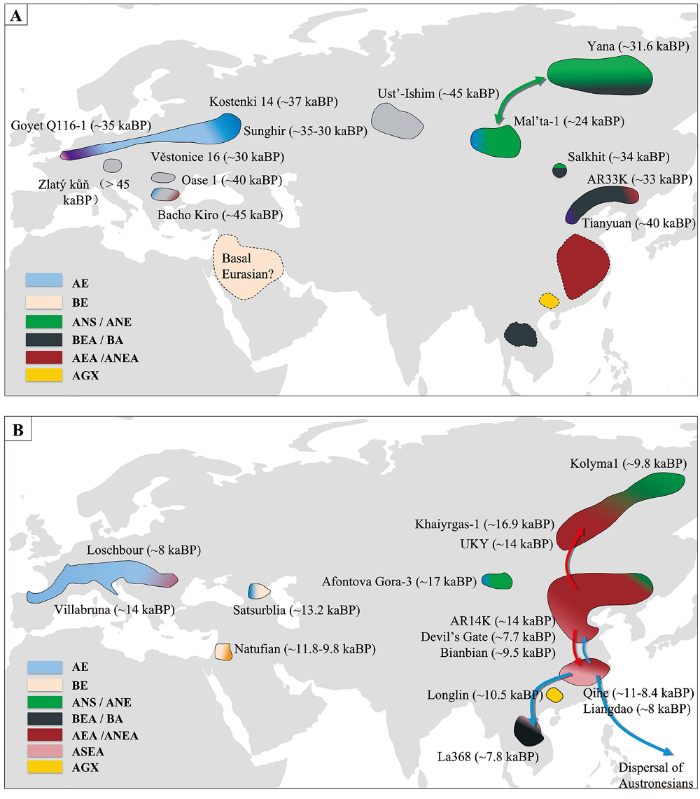
图1 45~10 kaBP欧亚大陆人群示意图
A)阶段1 Period I(45~19 kaBP);B)阶段2 Period II(19~10 kaBP):AE,以Kostenki 14为代表的古欧洲人群相关的遗传成分ancestry related to Ancient Europeans (represent by Kostenki 14);BE,与未经取样的古欧亚人群相关且在现今欧洲人群、部分古代和现今中东人群中可找到的遗传成分ancestry related to an unsampled population known as Basal Eurasians and found in small numbers in ancient and present-day populations of the Near East and in present-day Europeans;ANS,以Yana为代表的古西伯利亚北部人群相关的遗传成分ancestry related to ancient North Siberians (represent by Yana); ANE, ancestry related to ancient North Eurasians (represent by Mal’ta-1 and Afontova Gora-3);ANE,以Mal’ta-1和Afontova Gora-3为代表的古北欧亚人群相关的遗传成分;BEA,以田园洞人和AR33K为代表的基础东亚人相关的遗传成分;BA,以La368为代表的古代东南亚狩猎采集人群相关祖源成分;AEA,东亚古北方人群和东亚古南方人群分离前的亚洲人群;ANEA,以AR14K、扁扁洞、Devil’s Gate等个体为代表的东亚古北方人群;ASEA,以奇和洞、亮岛个体为代表的东亚古南方人群;AGX,以隆林个体为代表的古广西人群。图1A中的紫色代表田园洞人和Goyet Q116-1个体相联系的成分。虚线区域是指该遗传成分源自根据遗传数据所推测的人群状态,但未发现这一时期的古代样本。各颜色大致显示为某代表区域之间或之内的遗传成分组合,且颜色梯度显示为不同人群之间可能存在的遗传联系(如基因流)。
Fig.1 Schematic of Populations in Eurasia from 45 to 10 kaBP
AE, ; BE, ; ANS, ; BEA, ancestry related to Basal East Asians (represent by Tianyuan and AR33K); BA, ancestry related to Basal Asians (represented by La368); AEA, ancestry related to Ancient East Asians; ANEA, ancestry related to Ancient Northern East Asians (represented by AR14K, Bianbian, Devil’s Gate, etc); ASEA, ancestry related to Ancient Southern East Asians (represented by Qihe and Liangdao); AGX, ancestry related to Ancient Guangxi population (represented by Longlin). The purple color in Fig.1A shows the connection between Tianyuan and Goyet Q116-1. Broken lines indicate no ancient genetic samples have been found for a population with the inferred ancestry. Colors loosely indicate genetic groupings between or within a region, with color gradients showing the connections (i.e., gene flow) that may exist between different ancient populations.
2.1 阶段1(45~19 kaBP)
欧亚大陆史前现代人的古基因组研究表明,在约40 kaBP前,欧亚大陆至少存在6个不同的现代人群体(图1),分别为:
同样,Oase 1个体与14 kaBP及更早期的古欧洲人群、现今东亚人群和现今美洲原住民人群均拥有相似数量的等位基因,即拥有相近的亲缘关系。这一结果表明Oase 1个体所代表人群在欧洲无延续至今的直接后裔,是一种对现今欧亚人群无实质遗传贡献的独特人群;而且由于携有6%~9%的尼安德特人遗传成分,Oase 1个体可能代表了与尼安德特人具有密切互动关系的一支欧洲早期现代人群体[7]。
Zlatý k?ň个体所代表的人群,同样未给之后的欧洲人和亚洲人贡献基因成分。与同时期的其他现代人个体相似,Zlatý k?ň个体携有约3%的尼安德特人成分,但其含有尼安德特人基因片段的平均长度长于Ust-Ishim个体,这说明发生的基因重组次数少,前者的尼安德特人成分混入时间短于后者。Zlatý k?ň个体所代表的人群也是最早定居欧亚大陆的人群之一[18]。
2)以发现于保加利亚的46~43 kaBP的Bacho Kiro洞的3个个体为代表的古人群。这一人群相比于晚些时间出现在欧亚大陆西部地区的人群,与东亚和美洲的现代和古代人群关系更近。这表明他们属于一个早前迁入欧洲、但此前没有发现基因证据的群体,而且至少有部分与这一群体相关的基因成分延续到了后期的欧亚大陆人群中[19]。
3)以发现于中国北京约40 kaBP的田园洞人[20⇓-22]为代表的古人群,可称之为“古东亚人”(Basal East Asians, BEA)。这一人群广泛分布在欧亚大陆东部地区,与稍晚一些生存在黑龙江流域的AR33K个体(约33.6 kaBP)属于同一人群。研究发现,田园洞人较之现今欧洲人群和古欧洲人群来说,与现今东亚人群和美洲原住民人群有更近的亲缘关系[22]。相较于其他美洲原住民,某些南美洲原住民人群,特别是苏鲁人(Suruí)和卡利吉亚纳人(Karitiana),显示与大洋洲人、安达曼昂格人和田园洞人均有遗传联系,这表明美洲原住民人群的祖先们可能分批进入美洲,因而亚洲人群对其有不同程度的遗传贡献[22,23]。这些联系也表明,田园洞人的遗传成分至少存在于欧亚东部人群的亚群之中,且以某种形式一直延续到欧洲殖民者进入美洲之前。
4)以发现于西伯利亚西部约36 kaBP的Kostenki 14个体和发现于比利时约35 kaBP的Goyet Q116-1个体[24,25]为代表的古人群,可称之为“古欧洲人”(Ancient Europeans, AE)。“古欧洲人”较之于其他欧亚人群而言,与现今欧洲人群的关系更为相近[24,25],这表明某些与现今欧洲人群有遗传联系的现代人群体至少在35 kaBP已经广泛分布于欧亚大陆西部[24](图1)。发现于俄罗斯约34 kaBP的Sunghir个体与Kostenki 14个体及约34~26 kaBP的捷克V?stonice 16个体均有遗传联系,这意味着Kostenki 14所代表人群,至少有一部分被与V?stonice 16有亲缘关系的人群所替代[26]。
综合田园洞人、Kostenki 14、Goyet Q116-1等个体的古基因组研究可以发现,在约40 kaBP,遗传学意义上的欧洲人群已在欧洲(比利时)出现,遗传学意义上的亚洲人群也已在亚洲(中国)出现[22]。这也表明欧亚人群的分化(即欧洲人群和亚洲人群的分离)很可能早于40 kaBP,这与依据遗传突变率所推导的这两个人群的分歧时间(40~20 kaBP前至80~40 kaBP)一致[27⇓-29]。然而,虽然田园洞人与Goyet Q116-1个体在地理上相隔万里,但他们之间却存在着遗传联系。此外,他们分别与现今亚洲人群和欧洲人群具有更近的遗传联系,但是他们所共有的等位基因数量比早期亚洲人群和欧洲人群之间以简单分离模式所预期的多[22],这种现象产生的原因可能与亚洲人群和欧洲人群在分离之后仍发生过基因交流相关[30⇓-32]。考虑到Goyet Q116-1个体和田园洞人这两个个体生存时间之早和地理距离之远,Yang等认为当时的东亚人可能并未直接与欧洲人发生基因交流,而可能是跟某一个与这两个个体代表人群均相关的未知人群都发生过基因交流,而这个未知人群可能是从尚未分化的古欧亚人群中的某一亚群演化而来[22]。近期的一项研究发现,保加利亚46~43 kaBP的Bacho Kiro古人群,将Goyet Q116-1个体和田园洞人代表的两个人群联系了起来[19]。
距今4万年以来至末次盛冰期前的现代人群体中有些群体独立存在,还有一些群体与距今4万年以前的古人群相联系,比较具有代表性的有以下5个群体:
2)以发现于蒙古国东北部约34 kaBP的Salkhit个体为代表的古人群。Salkhit个体属于欧亚大陆东西部人群分离后形成的人群,而且有欧亚大陆西部人群的遗传成分。Salkhit个体携带有约75%的与田园洞人相关的祖源成分(BEA相关祖源成分),以及约25%的与Yana人群相关的祖源成分(ANS相关祖源成分)[35]。
3)以发现于中国黑龙江流域约33.6 kaBP的AR33K个体为代表的古人群。AR33K个体与田园洞人有最近的亲缘关系,但是AR33K个体与Goyet Q116-1个体的关系并非像田园洞人与Goyet Q116-1个体一样相近。而且,相比于其他美洲原住民人群,AR33K个体也没有与南美洲的苏鲁人(Surui)拥有更近的关系。另外,AR33K个体中也没有发现与Yana人群相关的祖源成分。这表明东亚北部地区可能在这一时期已经形成复杂的人群结构。田园洞人、AR33K个体及Salkhit个体的古基因组研究共同表明,末次盛冰期前,与田园洞人相关的人群在东亚北部地区广泛分布(分布范围至少覆盖田园洞人代表的华北平原、AR33K代表的中国东北地区和Salkhit代表的蒙古国东北部地区),该人群在蒙古国与Yana相关人群产生了基因混合,而这一时期的黑龙江流域的人群则保持独立特征[5]。
5)以现有遗传数据推测出来的一个未知人群,可称之为古欧亚人群(Basal Eurasians,BE),出现在欧洲人群和亚洲人群分离之前[36]。现今欧洲人群携有这种古欧亚人群的遗传成分,而这种遗传成分的获得主要发生在农业文明进入欧洲以后,因而相较于农业文明以前的古欧洲人群(即14 kaBP及更早期的不携有古欧亚人群(BE)遗传成分的古欧洲人群)而言,现今欧洲人群总体上与其他现代人群的关系较远。Lazaridis等[37]研究发现,古欧亚人群(BE)遗传成分在约34~14 kaBP前的近东人中存在较高比例,在约13 kaBP前高加索地区的狩猎采集人群中也开始发现古欧亚人群(BE)的遗传成分。另外,该人群显示同时具有欧洲人群和近东黎凡特地区的纳图夫人群(Natufians)的遗传特征。欧洲西部的狩猎采集人群与欧洲东部的非洲以外人群(non-African)至少在40 kaBP前就已经分离,而古欧亚人群的分歧时间更早,可能在非洲以外人群与灭绝古人类发生基因混合之前就已经分离[36,37],因此可能不具有或仅具有极低含量的尼安德特人成分[37]。古欧亚人群形成的时间相对较早,存在的时间可能比较长,且持续到更晚的时期,对欧洲和近东地区的现代人产生重要的遗传影响。现今欧洲人群所携有的尼安德特人成分低于现今欧亚地区的其他现代人[9,37⇓-39],很可能是因为古欧亚人群后期进入欧洲,导致欧洲人的尼安德特人成分被稀释。
2.2 阶段2 (19~10 kaBP)
末次盛冰期结束后,欧亚大陆各地区的现代人进入快速发展期。这一时期有些现代人群体与现今现代人群体之间存在一定的遗传连续性,也正是这一时期不同地域古人群的扩增、迁徙、融合等活动,逐渐奠定了现今现代人的遗传结构和分布格局。
在欧亚大陆东北部地区,末次盛冰期前后的人群不连续,但末次盛冰期结束后的人群则存在连续性。发现于中国黑龙江流域19 kaBP的AR19K个体相比于末次盛冰期前的古东亚人(如田园洞人和AR33K)而言,与末次盛冰期后至全新世的人群具有更大的遗传相似性,这表明在末次盛冰期即将结束时,东亚东北部的人群结构已经产生了颠覆性改变[5],这一情况不同于贝加尔湖区域人群在末次盛冰期前后的连续性[24,34]。相比于东亚古南方人群(Ancient Southern East Asians, ASEA;以中国福建约8.4 kaBP的奇和洞个体和台湾海峡约8.3 kaBP的亮岛个体为代表),AR19K个体与东亚古北方人群(Ancient Northern East Asians, ANEA;以中国山东9.5~7.7 kaBP的个体为代表)遗传关系更近。这表明在末次盛冰期结束时,东亚南北方古人群的遗传差异已经形成,而AR19K个体所代表的人群也被认为是最早期的东亚古北方人群(ANEA)[5]。这一人群后期以黑龙江流域的一系列时间连续的个体(AR14K等)、俄罗斯滨海地区(鬼门洞Devil’s Gate)约7.7 kaBP的个体、中国山东9.5~7.7 kaBP的个体(如扁扁洞Bianbian)、贝加尔湖区域约7.1~6.3 kaBP的个体作为代表[2,5,13,40]。自14 kaBP以来,这一人群在中国黑龙江流域具有遗传连续性,现今这一区域的一些少数民族仍然携有这些人群延续下来的遗传成分[40]。
这一时期分布在西伯利亚的古人群显示出不同祖源成分的混合,他们对后期出现的人群产生了重要影响。该人群被称之为古西伯利亚人群(Ancient Paleo-Siberians, APS),以发现于俄罗斯贝加尔湖地区约14 kaBP的UKY个体、远东地区约9.8 kaBP的Kolyma1个体为代表。这一人群以亚洲祖源成分为主,但混有古西伯利亚北部人群(ANS)相关的祖源成分,且Kolyma1代表着在美洲之外与美洲原住民人关系最近的人群[2,42]。在俄罗斯西伯利亚发现的约17 kaBP的Khaiyrgas-1个体所代表的群体与古西伯利亚人群(APS)具有十分相近的遗传关系[41],是该人群的早期代表之一。研究表明古西伯利亚人群(APS)的东亚祖源成分的最近来源便是14 kaBP以来的东亚古北方人群(ANEA)[5],这表明古西伯利亚人群(APS)可能与东亚古北方人群(ANEA)发生过基因混合。而古西伯利亚人群(APS)相关祖源成分也在山东博山(Boshan)、鬼门洞(Devil’s Gate)、贝加尔湖个体中存在,这表明至少有部分遗传成分从古西伯利亚人群(APS)混入到东亚古北方人群(ANEA)中,说明至少在8 kaBP前,古西伯利亚人群(APS)已经与东亚古北方人群(ANEA)有所联系[5,13,43]。
除与现今现代人有遗传联系的古人群外,东亚南部还发现了未对现今现代人产生遗传影响的史前现代人群体。发现于广西隆林(Longlin)约10.5 kaBP的个体所代表的古广西人群(Ancient Guangxi population, AGX),与此前发布的中国南方沿海的东亚古南方人群(ASEA,以此前福建个体为代表)及东南亚狩猎采集人群相关祖源人群(基础亚洲人Basal Asians, BA;以发现于老挝约8 kaBP的La368个体和马来西亚约4.3 kaBP的Ma911个体为代表的Hòabìnhian狩猎采集人群)[44]不同,属于此前未知的东亚现代人群体,对现今现代人未产生遗传影响。这一人群与东亚南北方祖源人群(ASEA和ANEA)具有很深分歧,并且至少持续到6 kaBP。而约9~6 kaBP的广西人群混合有当地祖源成分(AGX)、东亚古南方祖源成分(ASEA)及东南亚狩猎采集人群相关祖源成分(BA)[44],这表明了农业在这一区域兴起之前,人群已经有了广泛的混合,也说明在东亚和东南亚存在的3个独立祖源人群在史前时期互动频繁。
这一时期的欧亚大陆西部人群相比于更古老的欧洲人群,显示出与现今亚洲人群和近东人群更近的亲缘关系。约14 kaBP以来的欧洲中西部个体与一些跨越旧石器和新石器时代的文化个体(如意大利的Villabruna个体)密切相关,且显示出与现今近东人群和东亚人群有遗传联系[22,24,34]。高加索地区约13~10 kaBP的个体(如格鲁吉亚的Satsurblia和Kotias)与欧亚西部个体有较近的亲缘关系,且含有古欧亚人群(BE)的遗传成分。古欧亚人群(BE)的遗传成分在现今欧洲人群和近东人群中均有存在[24,36,37],尤其在近东人群中的含量最高,推算在旧石器时代晚期约12~9.8 kaBP的黎凡特地区,纳图夫人(Natufian)个体中古欧亚人群(BE)遗传成分的含量高达44%;在中石器时代文化约9.1~8.6 kaBP的伊朗个体(如伊朗的Hotu个体)中古欧亚人群(BE)遗传成分含量高达66%[37]。
除以上列举的主要史前现代人群体外,科学家们对全新世以来的更多材料进行了古基因组学研究,发现不同人群之间产生了多次交互影响的事件,对世界现代人群体的分布起到重要的影响。如:在贝加尔湖区域,新石器早期古北欧亚人(ANE)祖源人群,被与现今东亚北部人群相关的祖源人群大量替代[45]。进入全新世之后,东亚南北方人群频繁互动,但整体而言主要有三次大规模的南向基因流动,包括:1)东亚古北方人群相关的遗传成分在东亚南方人群中有所增加;2)具有东亚遗传成分的人群向东南亚扩散,并与基础亚洲人(BA)混合;3)东亚古南方人群祖源成分,通过南岛语系人群向东南亚岛屿和太平洋岛屿扩散[13,43]。人群的互动为现今的东亚大陆人群遗传格局的形成奠定了基础。
3 展望
史前现代人群体的古基因组研究揭示了欧亚大陆丰富的人类历史。对这一时期的现代人展开的古基因组研究,已揭示出欧亚大陆人群丰富多彩的遗传演化历史,极大地提升了我们对史前人群的更替和迁徙历史等相关问题的理解和认识[48]。近年来,欧亚大陆东部的人类古基因组研究快速发展,尤其一些东亚南方样本基因组信息的成功获取,为我们发现和了解更多史前人类群体的遗传历史提供了重要支撑[13,47]。虽然欧亚大陆史前现代人基因混合和人群结构相关的许多问题已得到了答案,但随着研究的深入,新的科学问题也在不断出现,例如,灭绝古人类对不同区域的史前现代人和现今现代人群体的基因贡献程度及影响如何?古基因组是否能发现更多独立的史前现代人群体?史前现代人如何发展、演变、交流,并逐渐形成后期人群及现今人群格局的?
随着古DNA实验技术的日益成熟和分析方法的不断完善,研究者们也能够扩展除欧亚大陆以外的材料以进行研究,有助于更全面揭示早期现代人的群体结构,以及灭绝人类在现代人演化历史中所扮演角色等相关问题。近年来,科学家们开展了许多关于新石器时代以来现代人群的古基因组研究,尤其是东亚[5,13,47,49,50]、欧洲[8,35,36]和美洲[51]人群。古基因组研究团队在长时间尺度下开展人群遗传演化分析时需要尽量结合多个学科,从不同角度进行探讨,才能够更好地还原真实的人群演化图景。
https://www.anthropol.ac.cn/article/2023/1000-3193/1000-3193-42-03-412.shtml
 选择“Disable on www.wenxuecity.com”
选择“Disable on www.wenxuecity.com”
 选择“don't run on pages on this domain”
选择“don't run on pages on this domain”